
Descripció Física
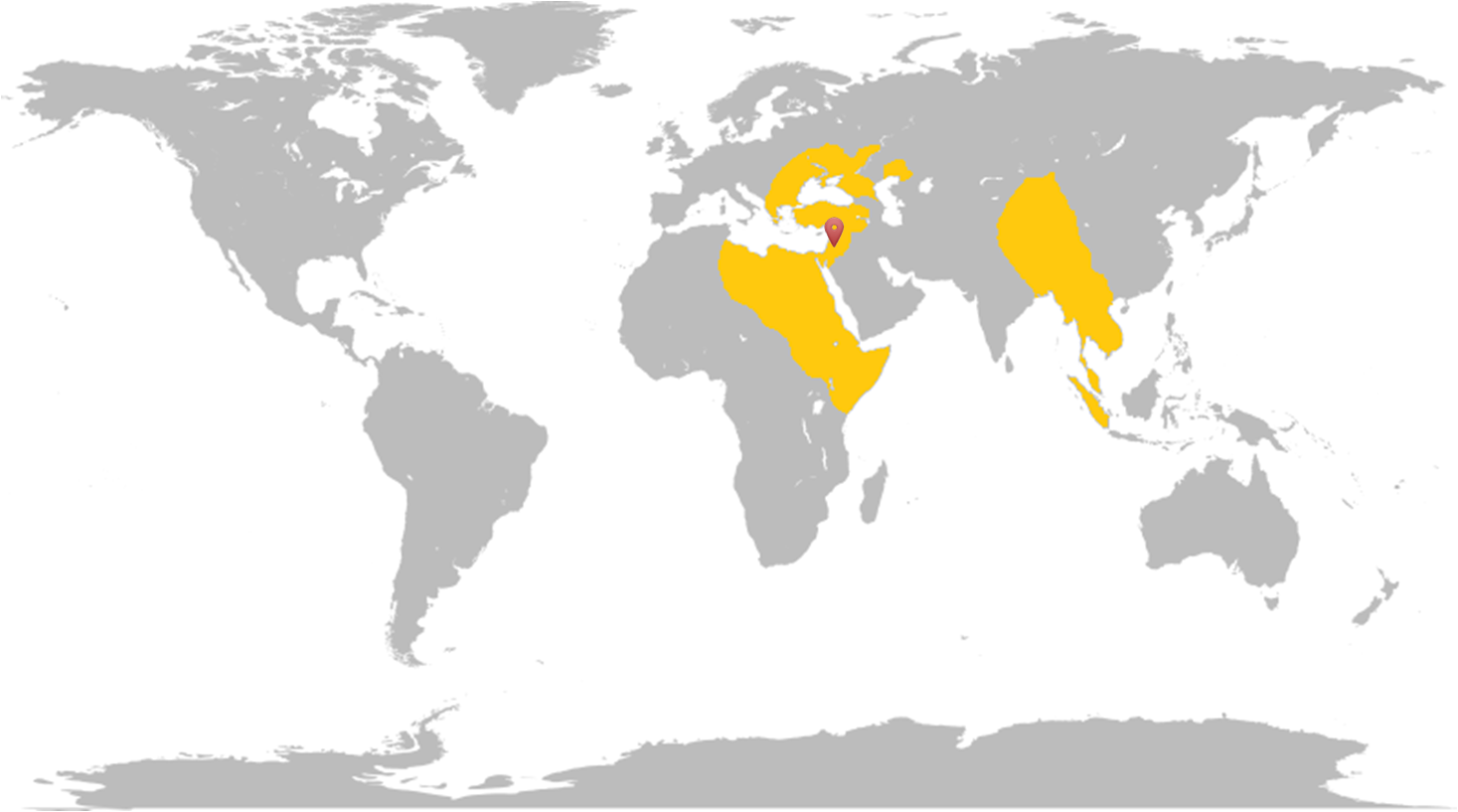
La rata talp cega de les muntanyes de Galilea o Nannospalax galili, forma part de la família Spalacidae subfamília Spalacinae i la superèspecie Spalax ehrenbergi, i es troba a Israel ![]() .
.
La família Spalacidae és un grup divers de rosegadors del vell món, adaptats a l’excavació i la vida subterrània. Consisteix en 36 espècies i 6 gèneres, distribuïts en quatre subfamílies: Myospalacinae, Rhizomyinae, Spalacinae i Tachyoryctinae.
Es troben a Europa des d’Ucraïna i els Balcans fins al Mediterrani Oriental, a l’Àfrica des de Líbia per l’oest fins al nord de Tanzània i a l’Àsia des del sud de Sibèria fins a Sumatra i a l’oest de Xina.
Construeixen caus en pastures, matolls, àrees agrícoles i boscos. Eviten els deserts, ja que prefereixen sòls més humits. Alguns habiten regions muntanyoses, arribant als 4.000 metres d’altitud.
Els espalàcids tenen un cos robust i arrodonit, els ulls i les orelles externes reduïdes, la pell densa i les extremitats curtes. El seu cap és ample, tenen poderosos músculs al coll i els grans incisius es projecten davant dels llavis, excepte en els Myospalacines que excaven amb els avantbraços. La mida corporal varia des de 130 mil·límetres i 100 grams en Spalax leucodon, fins a 480 mil·límetres i 4 kg de Rhizomys sumatrensis. Pel que fa a Spalax ehrenbergi, es troba al voltant dels 200 mil·límetres. En algunes espècies, els macles són més grans i, en altres, no s’observa dimorfisme sexual.

Spalax ehrenbergi

Rhizomys sumatrensis

Spalax leucodon
Són animals solitaris, sedentaris i molt agressius. Són actius durant el dia i només algunes espècies migren estacionalment per escassetat de menjar. La major part de la seva vida es dóna sota terra, on construeixen elaborats sistemes de túnels cavant amb els seus incisius o urpes i empenyent la terra amb les potes del darrere. Aquestes galeries, inclouen àrees definides per a dormir, sortir a la superfície i defecar.
Pel fet de viure sota terra, els espalàcids no utilitzen el seu sistema de visió. Tot i així, en algunes espècies els ulls encara són fotoreceptius, fet que els permet distingir les diferents etapes del dia. En canvi, tenen molt desenvolupat el tacte, l’oïda i l’olfacte. Les feromones són una important forma de comunicació, i algunes espècies piquen amb el cap a les parets dels seus túnels i capten les vibracions produïdes per altres per transmetre informació. Diferenciem sis tipus de vocalitzacions: d’atac, de plor, d’invitació, de festeig, d’alliberament i d’amenaces.
La seva dieta consisteix en la ingesta d’arrels, bulbs, rizomes i altres parts subterrànies de les plantes. De tant en tant, algunes espècies poden menjar brots, fulles, llavors, fruites, insectes i altres artròpodes. Molts emmagatzemen grans quantitats d’aliments en els seus caus.
Per a la reproducció, mascles i femelles s’associen només durant el festeig i l’aparellament. Solen tenir una o dues ventrades per any i des d’1 a 5 nounats. El temps de gestació es troba entre les 4 i les 7 setmanes, varia entre espècies i depèn de la localització. Les femelles construeixen caus subterranis per tenir cura de les cries durant les primeres 4-6 setmanes, però les cries no els abandonen fins als 2-3 mesos.
Els principals predadors que els cacen quan surten a la superfície són els mussols, les serps, les àguiles i altres mamífers petits i, són parasitats per nematodes, àcars, paparres i puces.
De cara a l’ecosistema són importants perquè, a més de ser preses, la seva activitat excavadora afecta la distribució de nutrients, aire i aigua al sòl.
Torna a dalt
Torna a la Introducció
Descripció del Genoma
Spalax galili és un model excel·lent per a estudiar l’adaptació dels mamífers a la vida subterrània. Aquesta espècie passa tota la seva vida sota terra, protegint-se dels depredadors i de les fluctuacions climàtiques, mentre que s’enfronta a múltiples factors d’estrès com són la foscor, la hipòxia, la hipercàpnia i l’elevada patogenicitat.
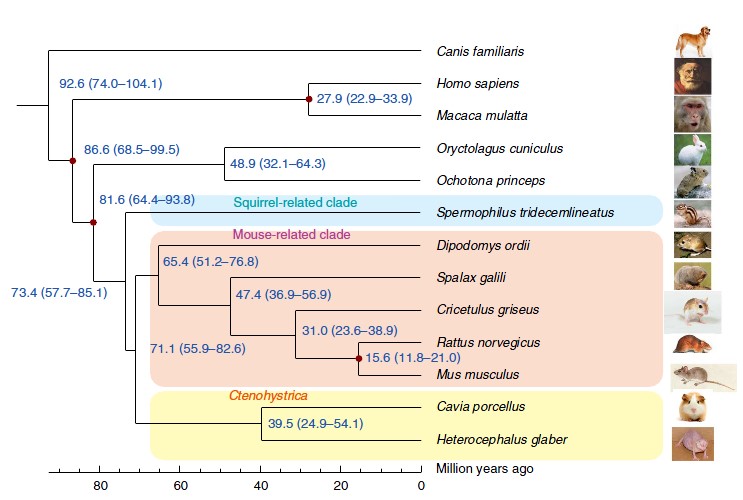
El genoma de la rata talp cega conté 22.168 gens codificants i mostra la major similitud amb el hàmster xinès (Cricetulus griseus). Es troba dins del grup dels rosegadors (Muroidea) i va divergir de les rates, els ratolins i el hàmster xinès fa 47 milions d’anys. Mitjançant la comparació amb altres espècies s’ha estimat que 139 famílies de gens s’han expandit i 50 s’han contret. Així, s’han detectat 39 gens perduts i 259 pseudogens, i també 48 gens seleccionats.
L’anàlisi del transcriptoma de la placenta, ha portat a la conclusió que la divergència entre la rata talp nua (Heterocephalus glaber) i la rata talp cega es va donar abans de l’origen dels Muroidea.
Diferents característiques morfològiques (absència de cua i coll, cap ample i aplanat i orella externa reduïda) reflecteixen l’adaptació al moviment dins de l’estructura de túnels. De la mateixa manera, les urpes els incisius i el sistema muscular estan adaptats a l’excavació. Addicionalment, la falta de llum ha portat a un mosaic de trets complexos respecte la percepció de la llum, incloent una gran regressió ocular fins a un ull minúscul i subcutani. Dels 259 pseudogens trobats en el genoma de la rata talp cega, 22 estan involucrats en el sistema de visió.
La comparació amb la rata talp nua, ha revelat una evolució convergent dels gens que regulen el cicle circardià (Clock) responsables de l’adaptació a la foscor. Les proteïnes Clock han expandit la seva regió rica en glutamina, involucrada en la ritmicitat circardiana, i han canviat la seva composició d’aminoàcids.
A més, s’ha observat que les rates talp cegues tenen un volum cerebral major respecte a altres rosegadors amb mida corporal similar. El gen SEC23a, crucial per la morfogènesi del crani i la cara en humans, mostra una forma de splicing alternativa en la rata talp cega. I la família de factors neurotròfics derivats de línies cel·lulars glials s’ha expandit, degut a les múltiples duplicacions del gen Gfra1 que és un factor de supervivència neuronal. La seva sobrerepresentació pot estar relacionada amb el creixement del neocòrtex causat pel desenvolupament dels sistemes d’orientació per vibracions, tàctils, vocals, olfactoris i magnètics i per l’ampliació de les estructures motores relacionades amb l’excavació.
En el genoma de Spalax galili s’ha identificat una reducció en els reordenaments cromosòmics, una representació elevada de elements SINE i una alta taxa d’edició del RNA/DNA.
En total, conté 18 reordenaments intercromosòmics, dels quals 17 es van donar en el llinatge Muridae i només 1 en el llinatge de la rata talp cega. Les dades indiquen una evolució del genoma amb una taxa de reordenaments del 0,56% després de la separació amb els ratolins (fa uns 47,6 milions d’anys).
Un 43,9% són regions repetitives, de les quals els més prevalents són els retrotransposons (32,5%). La proporció d’elements SINE (11,8%) és molt similar a la dels humans (13,7%) i més alta que en la majoria de rosegadors (6,5% en ratolins, 5,6% en rates). D’aquests, els més abundants són B1, B2 i B4, que són elements induïbles per factors d’estrès. Per tant, la seva expressió elevada està molt probablement relacionada amb l’exposició a la hipòxia.
El genoma de la rata talp cega conté un gen APOBEC3 (A3), el producte del qual pot introduir sèries de mutacions C a U a la cadena negativa d’un retroelement que s’està replicant i la seva preferència són els motiu GxA dels retroelements LTR (long term repeats). L’abundància de SINE afavoreix la formació d’estructures de RNA en doble cadena, que són el substrat preferit de l’enzim ADAR i, per tant, augmenten la taxa d’edició de RNA.
Per tal de sobreviure i dur a terme activitats intenses en un medi hipòxic i hipercàpnic amb fluctuacions dels nivells d’oxigen, mostren diferents adaptacions: millora de la funció i l’aportació d’oxigen al miocardi, freqüències cardíaques i respiratòries adaptatives, concentracions d’hemoglobina i hematòcrit elevades, ampliació de la superfície alveolar i el volum de capil·lars i un increment de les mitocòndries i els capil·lars en els teixits.
Un gran nombre de gens estan implicats en la tolerància a la hipòxia. Així, trobem una expressió constitutiva i elevada de mRNAs i proteïnes d’hemoglobina, mioglobina, haptoglobina, neuroglobina i citoglobina. També s’ha observat una major expressió de gens relacionats amb la defensa cel·lular, la resposta a estímuls de l’hormona esteroidea, biogènesi ribosomal i mitocondrial, splicing de RNA, citoesquelet i regulació de la circulació i la pressió sanguínia, la gana i els ritmes circardians.
Una adaptació als nivells elevats de diòxid de carboni podria ser la que es troba en Scn9a, que codifica per un nociceptor en forma canal de sodi (Nav1.7) i mostra convergència evolutiva amb al rata talp nua. En el domini IV de la proteïna Nav1.7, el motiu altament conservat d’aminoàcids amb càrrega positiva, KKV, s’ha substituït pel motiu amb carrega negativa EKD en la rata talp cega i EKE en la rata tap nua. Mitjançant l’atracció de protons podria protegir els teixits dels danys induïts per l’acidosi als teixits.
A més, també s’ha observat un canvi d’aminoàcid en la regió d’unió al DNA de la proteïna p53 (gen Tp53), que correspon a una mutació associada a tumors en humans. Amb aquesta mutació, la proteïna perd la capacitat d’induir l’apoptosi en favor de l’aturada del cicle cel·lular i l’activació d’elements reparadors del DNA, per tal d’evitar l’excessiva pèrdua de cèl·lules davant la hipòxia.
La falta de control per part de p53 porta a la transcripció massiva dels SINE B1 i B2, acompanyada de l’activació de la resposta d’interferó (TRAIN). Les cèl·lules pre-malignes secreten interferó, que provoca la necrosi massiva de les cèl·lules veïnes. El gen IfnbI, que codifica per l’interferó-β1, es troba duplicat en el genoma de la rata talp cega i els gens Mx1, que formen part de la via de senyalització del’ interferó-β1, i altres gens involucrats en la regulació de la mort cel·lular i la inflamació (Nfkb, Tnfrsf1a, Birc3, Fem1b i Aifm1), mostren una expansió. A més, gens involucrats en la necrosi i la inflamació (Tnfrsf1a, Tnfsf15 i Nfkb1) mostren selecció positiva. Per tant,el mecanisme que explica la resistència al càncer tant espontani com induït per factors químics, molt probablement està basat en la major resposta inflamatòria via amplificació gènica de la via de l’interferó-β1.
D’altra banda, també s’ha observat que expressen una forma alternativa d’splicing de l’heparanasa, que actua com a dominant negatiu per reprimir la degradació d’heparan sulfat en la matriu extracel·lular i, per tant, actua com a factor antitumoral.
També es possible que l’àcid hialurònic contribueixi a la longevitat d’aquesta espècie com en el cas de la rata talp nua. En la rata talp cega, també predomina la forma d’elevada massa molecular que té propietats antiinflamatòries, ja que s’uneix a CD44 i atura el cicle cel·lular reprimint les senyals mitogèniques. A més, aquesta molècula presenta un efecte antioxidant i podria millorar els nivells d’estrès oxidatiu.